First published in: Gajah 28 (2008), pp. 44-52
Elephant Cognition: A Review of Recent Experiments
Moti Nissani
Department of Biological Sciences, Wayne State University, Detroit, Michigan, USA
One of the greatest remaining biological mysteries, says Marian Dawkins, is “what goes on inside non-human minds” (1993). Indeed, the question whether some animals, at least, think, it could be argued, has fundamental implications to our world view and to the way we interact with animals at homes, farms, parks, zoos, and elsewhere.
This brief review focuses on the question whether elephants—one contender for high intelligence in animals—can act thoughtfully. For the purposes of the present discussion, we shall arbitrarily narrow down the meaning of thinking to just one aspect: the ability to plan future actions in one’s head, before embarking on any specific course of action.
Discrimination experiments
Most of the evidence in this section is based on the pioneering work of Rensch (1957) and Altevogt (1990) with a single young Asian female at the Münster Zoo, and on our own work with 22 Asians at the Detroit Zoological Institute and Burma. In Burma, elephants first learned to remove a lid from a bucket, or to displace a box covering a hole in the ground, in order to obtain a desirable food item (Nissani et al. 2005; Nissani 2006). This was followed by either a black/white discrimination task, a large/small transposition task, or the placement of either box or lid on the ground so that they no longer obstructed access to the food, and then observing the elephant’s reaction. For the most part, these experiments followed Fabre’s (1915) and Thorndike’s (1911) protocols of distinguishing trial-and-error learning from thoughtful behavior.
Laborious, gradual, process of learning
Thorndike, (1911, p. 73) argued that his cats and dogs escaped puzzle boxes through a mindless process of trial and error. Because understanding something as simple as pulling a loop to open a door must occur rapidly or not at all, it should have induced, at some point during the repeated introductions of his animals into the box, a sudden reduction in escape time. The actual, gradual, slope of the time-curve that he did observe suggested to him that his subjects failed to understand the cause-effect relationships between their actions and escape.
Like Thorndike’s cats and dogs, all 13 Burmese elephants that mastered our black/white or large/ small discrimination tasks did so gradually, with the number of correct responses rising by fits and starts, over several sessions, from chance level to near-perfect performance (Fig. 1).
The same logic applies to the pre-training of the elephants in our discrimination experiments, which involved learning to remove a lid from a bucket or to displace a box to uncover a hole in the ground. Here too, one never sees sudden improvements. On average, the 20 elephants taking part in these experiments required 3.4 sessions to learn the task, imperceptibly nearing the experimenter’s goal.
In comparison to some other animals, elephants’ performance is unremarkable
Rensch and Altevogt’s young Asian elephant needed 330 trials, over a period of several days, to consistently choose the reinforced response in her first discrimination task (Rensch 1957). In an experiment which employed another sense modality, an 8-year-old took 7.5 months to distinguish 12 tones (Reinert, cited in Altevogt 1990, p. 474).
Likewise, the 13 elephants in our sample, which acquired our easier black/white or large/small visual discrimination tasks (mean age=13.2 years) did so in an average of 3 sessions and 154 trials (see also the section on short-term memory, below).
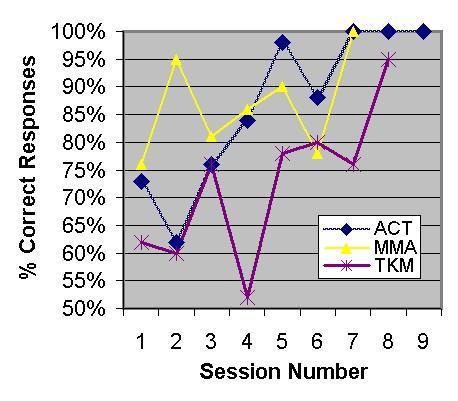 Figure 1. Learning curves for three representative elephants (ACT, MMA, TKM) in a white+/black- visual discrimination task.
Figure 1. Learning curves for three representative elephants (ACT, MMA, TKM) in a white+/black- visual discrimination task.
Additionaly, the remaining 7 Burmese elephants (mean age=29.3 years) failed to reach criterion in an average of 6.6 sessions and 332 trials, suggesting that some elephants may be either unable to acquire simple visual discriminations or that they require an inordinate number of trials to do so.
A acuird bhavioral sequence persists after it no longer serves a purpose
Thorndike (1911) reported that once an animal learned to escape from a puzzle box by performing a certain action (e.g. pulling a loop), on subsequent introductions to the box, and after the action no longer served a purpose (e.g., the cover of the box had been removed), the animal continued to perform that action. He argued that this supported the notion that the animal solved the task mechanically, without understanding the causal link between action and consequence.
Similarly, four logging elephants were trained to remove food from a coverless bucket by inserting their trunk into the bucket (Nissani 2006). They were next trained to remove a lid from the top of that bucket to obtain food. Once this behavioral sequence was established, the lid was placed alongside the bucket so that it no longer obstructed access to the food. If the elephants understood the nature of the task, they might be expected to ignore the side lid in the first few trials and retrieve the reward directly, as they used to do before the lid was introduced. On the other hand, if they did not understand what they were doing, in the first few trials they might continue to remove the lid before inserting their trunk into the coverless bucket and retrieving the food. Observations accorded entirely with the mindless learning hypothesis: in the fi rst 5 critical trials, when the lid was placed alongside the bucket and no longer obstructed access to the reward, each elephant continued to remove the lid before retrieving the reward. A number of variations on this basic design involving 11 additional elephants, and numerous controls, amply confirmed this conclusion. It appears probable, therefore, that when acquiring the obstacle-removal task, elephants respond to the temporal contiguity of the two events, not to their causal relationship.
Long-term memory
Learning to discriminate between 20 pairs of symbols, a young elephant is said to have performed superbly on a test that combined all 20. The test lasted several hours, yet her performance actually improved toward the end. A year later, her scores ranged from 63 to 100 percent (Rensch 1957). After a break of more than 32 years, when she was about 40, the elephant remembered the experimental situation and sequence, but could no longer reliably choose the 20 correct symbols (Altevogt 1990).
Likewise, one of three elephants that learned a simple discrimination task remembered it eight years later (reported in Stevens 1978). More convincingly, the single elephant that learned to distinguish 12 pure tones was able to distinguish 11 sounds after 19 weeks and 9 sounds after an additional interval of 1.5 years (Reinert, cited in Altevogt 1990).
Short-term memory
Four circus elephants had to choose one of five identical boxes in which a morsel of food was placed before their eyes, yielding a 76% correct response rate as the baseline. In delayed response experiments, the best performing animal scored at chance level after a 15-second delay. When the choice was limited to one of three identical boards, chance level was reached after a 45second delay (Grzimek 1944).
Our investigations of short-term memory of Burmese logging elephants yielded similar results. Two experimenters stood 1 m apart, facing the elephant at a 45° angle to its right and left, each holding a bucket. To help the elephant remember, the buckets differed in size and brightness. In each trial, a third experimenter, standing at a distance of some 2 m from the bucket-holders and directly facing the elephant, tossed a 2-4 cm piece of sugarcane into one of the buckets (for visual details see Nissani & Hoefl er-Nissani 2004). At pre-training 1, the two buckets rested on the ground. At pre-training 2, the buckets were placed on the ground right after the sugarcane loudly hit the bottom of one of the buckets, in full sight of the elephant, who now had to choose the correct bucket in order to secure the food reward. After the elephant mastered this second pre-training task, in experimental trials, following the toss and sugarcane entry into one bucket, the bucket holders placed the buckets behind their backs, hidden from the elephant, and kept them there for a specified period of time. Both holders then simultaneously placed the buckets on the ground in front of them, within easy reach of the trunk. Various control measures were employed to rule out reliance on experimenter bias, smells, sights, or sounds, in choosing the bucket. Interobserver reliability was measured by comparing the written records of one experimenter to a video record maintained by another.
Of the six elephants taking part in this experiment, one did not meet predetermined criteria for the absence of experimenter bias, and two others failed to reach pre-training criterion (in 8 and 18 sessions, respectively). The other three elephants reached that criterion by the 3rd or 4th session and their performance in experimental sessions improved over time. In their last session, when the buckets were lowered to the ground after a 4 seconds delay, the three elephants retrieved food from the correct bucket in 80-100% of the trials. When the duration of the waiting interval was increased to 6 seconds and was accompanied by an interference (walking towards the elephant and giving it a piece of sugarcane), the percentage of correct responses declined to 60%-67%. When the duration or level of difficulty of the task were raised, subjects’ performance declined still further.
We may note in passing that the two experiments described in this section strikingly differ from short-term tests with human beings. Tests with humans are designed (by using nonsense syllables, for instance) to prevent the formation of mnemonic rules. In the few cases where subjects manage to develop such rules, extraordinary feats of memory are observed. By contrast, the two elephant protocols above made no effort to forestall the formation of mnemonic rules; had these same protocols been applied to human subjects, many such subjects might have developed such rules as “move forward the front leg on the side of the appropriate bucket and, once the bucket is lowered, select the bucket nearer this leg.”
Williams (1950) says: “I don’t believe that an ‘elephant never forgets,’ but I should scarcely be surprised if he tied a knot in his trunk to remember something, if he wanted to.” Our elephants appeared to enjoy the activities and the unusual treats they were receiving, and one might expect that Grzimek’s elephants did too, but not one of the nine seemed capable of maximizing food intake by developing a simple memory-enhancing strategy. If elephants think, either they think at a lower level than this, or their thinking is qualitatively different from ours.
Mirror self-referential behavior
A 1989 traditional mark test of two Asian elephants failed to elicit self-referential response to experimentally-induced visible alterations in their body image. Because this failure could potentially be ascribed to the poor vision of elephants (Povinelli 1989), another study (Fig. 2) of two captive Asian elephants at the Detroit Zoological Institute bypassed the traditional painted mark test, relying instead on the elephants’ response to a more visually conspicuous object (a white turkey feather taped to their forehead). Despite the alternative design, neither subject engaged in mirror self-referential behavior (Nissani & Hoefler-Nissani 2006). However, another Asian elephant did pass the mark test (Simonet 2000; Simonet et al. 2000) and, more recently, Plotnik et al. (2006) reported that one of three Asian females at the Bronx Zoo passed the mark test as well.

We do not yet know whether mirror self-referential behavior implies a self-concept. Likewise, we cannot yet be sure whether elephants are capable of self-referential behavior. In chimpanzees, where such behavior has been repeatedly observed (Swartz & Evans 1991; Anderson 1996), not all individuals show this capacity, so the conflicting results described here could be ascribed to the fact that such behavior is only manifested by some individuals of a species but not by others.
Applying the bird string-drawing paradigm to elephants
In 1956, Thorpe commented: “The ability to pull up food which is suspended by a thread, the pulled in loop being held by the foot while the bird reaches with its beak for the next pull, is doubtfully inborn and it has been subject to many experiments. The act appears at fi rst sight to be a real and sudden solution of the problem from the start, and thus to qualify for inclusion under “insight learning” (Thorpe 1956, p. 333). Successful performance in this task has been documented in well over ten bird species.
By using a retractable (bungee) cord, we were able to apply the string-drawing paradigm to the two elephants of the Detroit Zoological Institute (Nissani 2004). Both mastered the problem, but, although insight could not be ruled out, our observations were more consistent with a trial-and-error acquisition of the task. The two elephants acquired the behavior gradually, by fits and starts, and seemed unable to transfer their skill across a change in physical stimuli (Mackintosh et al. 1985), e.g., successfully retrieving the reward when the string was tied to a pole on the elephant’s side of the cord (Nissani 2004).
To cast additional light on this application of the string-drawing paradigm to elephants, we presented seven Burmese elephants with a retractable cord. Here, elephants were first presented with a tamarind or salt-laced 80 cm simple, non-retractable rope to whose end a 2030 cm piece of sugarcane was loosely attached (pre-training 1). Next (pre-training 2), a longer simple rope was used. Next (pre-training 3), the retractable cord later employed in the experiment itself was used, but without being tied at its end to a heavy log, so that, in this third pre-training phase, the cord functioned as a simple rope. The experiment itself involved tying the retractable cord to a heavy log a few meters away from the elephant, so that the sugarcane tied close to the cord’s end could only be retrieved by repeated, coordinated, action of the trunk and another body part.
At the end of pre-training, which lasted 1-3 sessions, all elephants were able to effortlessly pull a long rope, in a series of 4-7 pulls, to obtain a piece of sugarcane tied to its end.
All seven logging elephants fully mastered the string-drawing sequence within 1-3 experimental sessions. In all cases of retractable rope pulling, the sequence involved pulling by the trunk, and then securing the rope by either foot or mouth. After 2-6 coordinated pulls, while still holding the rope with either mouth or foot, the elephants disengaged the sugarcane from the rope while still using mouth or forefoot as an anchor, and then consumed the sugarcane. All elephants seemed to be flexible about the use of anchor, interchangeably using mouth, foreleg, or both.
To test the transferability of this skill, two of these elephants were taken to a bridge. After a few trials of simple rope pulling, a long simple rope with a heavy rock and a piece of sugarcane at one end was tied by its other end to a rail and then thrown over the bridge. One elephant secured the rope immediately, in 5 coordinated trunk/mouth draws. The second elephant, in her fi rst trial, used forelegs and mouth as anchors, as she did in the earlier retractable cord variation, but close to the end of the first trial, wrapped the cord around her trunk. On the second and subsequent trials, she only wrapped the rope around her trunk until both the sugarcane and the accompanying heavy rock landed on the bridge.
It remains to be seen whether something like retractable rope pulling takes place naturally in the wild. For instance, do elephants take a branch down with their trunk, keep it in place with their foot, and munch on the leaves? On a visit to a remote logging camp in the forest, I did see behavior equivalent to wrapping a rope around the trunk. One logging elephant grabbed the end of a long (at least 5 m) creeping edible vine wrapped around a tall tree, a vine which looked like a thin rope, disengaged it from the tree by wrapping it around its trunk 4-5 times, and consumed it.
To sum up this series of string-drawing experiments in elephants, we can say that all nine Asian elephants we tested in Michigan and Burma mastered this task. We cannot however decide whether the awkwardness they showed on mastering the task was traceable to the novelty of the stimuli that surrounded this task or to a lack of understanding of its nature. Likewise, one elephant failed a simple transferability test (Nissani 2004) while two others passed the bridge transferability test. It is possible, but not yet proven, that elephants perform actions akin to pulling/anchoring in the wild, and that the conceptual aspects of the task presented no novelty for them. More ingenious experimental designs than the ones presented here are needed before we can prove or disprove the existence of insight in the string-drawing paradigm in either birds or elephants.
Competitive food procurement
At an earlier experiment at the Detroit Zoological Institute (Nissani 2004), we observed two elephants immediately securing objects from one of the two openings of an inflexible tube, 5 cm in diameter, following the placement of a small food item in that tube. They expertly placed their trunk tightly over one of the tube’s openings and either sucked the object toward them and ate it, or blew it out and retrieved it when it fell to the ground. We next applied their proficiency in this task to a competitive food procurement task.
When singly confronted with the tube, both elephants either sucked or blew, suggesting a random retrieving strategy. However, when both elephants were placed in a competitive situation in which sucking would have landed the bagel fragment or sugar cube in one’s mouth, while blowing would end it on the ground near its companion, from the very start both elephants almost always sucked the morsel, a reward-maximizing behavior consistent with the view that they understood the situation. Many additional variations of this setup (Nissani 2004) showed that both elephants were capable of adjusting their behavior to answer the logical demands of the task.
We have since subjected these intriguing results to two additional test implications. Upon first being presented with the tube, the two Detroit elephants were immediately able to expertly obtain the food, thus raising the possibility that this was a variation of a familiar task, which they were trained to perform earlier in life. To throw some light on this possibility, we have tested the ability of two Burmese females, 16 and 17 years of age, to retrieve food from an identical tube. They both tried repeatedly, in two daily sessions lasting more than one hour each, to obtain the food, by wrapping their trunk around the length of the tube, tilting the tube, or using force. Their failure to apply either sucking or blowing is consistent with (but does not prove) the notion that, prior to our experiments with the two Detroit elephants, they had been trained in a similar task.
We also subjected the two Detroit elephants to a transferability test (Fig. 3). We used a shorter and wider inflexible PVC pipe, 45 cm in length and 15 cm in diameter. Both elephants could comfortably insert their trunk all the way through this pipe. The pipe itself was kept 1 m horizontally above ground, tied securely to a heavy stool. We placed about a third of a bagel inside the tube, and, to forestall its suction inside the tube, wedged it between two snugly-fitting round pieces of wood.
Owing to their poor visual acuity, both elephants could not see the food once it dropped to the ground, and had to grope for it with their trunk. To overcome this problem, a big pan was placed on the ground at the end of each tube, so that the elephants only had to search for the food inside the pan.
We now placed our two elephants in a competitive situation conceptually similar to the one they faced earlier in the sucking and blowing task. If they understood the requirements of the task, they might be expected to enter the tube on their partner’s side and push the food towards themselves. If they did not understand, the pushing direction, in the first few trials at least, should remain random.

The data from Wanda are subject to two reservations. First, she was standing close to the dominant Winky, and her behavioral options were therefore constrained. Second, owing to earlier injuries, she was unable to perform certain actions with her trunk.
In the first competitive session, the more dominant Winky pushed the bagel towards Wanda on the first 6 trials; Wanda then consumed the bagel, leading Winky to twice raise her trunk in a threatening gesture. Then, on the 7th trial, Winky pushed the bagel towards herself and ate it. On the last trial, she pushed it again towards Wanda, and Wanda consumed it. Wanda pushed the bagel only once, towards Winky, who consumed it.
The second session consisted of 33 consecutive trials, of which 30 yielded clear results. The session and the entire experiment had to discontinue after trial 33 owing to the increasing intensity of Winky’s agonistic behavior towards Wanda. In these 30 trials, Wanda pushed the bagel towards herself and received the reward in 5 trials. She pushed the bagel towards Winky, failing to get the reward, in 12 trials. Winky pushed the bagel towards herself and ate it in 7 trials. She pushed it towards Wanda in 6 trials, failing to get it in 3, and getting it in 3 (by retrieving it from Wanda’s pan).
On one trial in that last session, each elephant simultaneously pushed from her side at the same time, each counterproductively struggling against the other (the intelligent thing to do would have been to let the other push the reward towards oneself, waiting for it to be dropped on one’s side, and then picking it up quickly).
This informal, preliminary, investigation suggests that the two elephants did not understand the nature of this competitive task. Since this task was conceptually similar to the sucking and blowing competitive task described earlier, their failure casts doubts on our earlier insight interpretation of that task.
Do elephants know that people see? (The Povinelli Paradigm)
In a series of experiments, six young chimpanzees faced one trainer who could see them and one who could not (Povinelli & Eddy 1996). Startlingly, in the first few trials, in all but one variation, the apes consistently performed at chance level. For instance, they were as likely to beg food from a trainer with a bucket fully covering her head as from a trainer with a bucket over her shoulder.
A preliminary application of Povinelli’s protocol to the two elephants of the Detroit Zoological Institute (Fig. 4), and a more complete experiment with six chimpanzees of that Institute, attempted to address methodological limitations of Povinelli et al.’s work (for a discussion, see Nissani 2004). In these experiments, both species performed significantly above chance in some conditions (Nissani 2004).
Extensive additional experiments with 14 logging elephants in two geographically isolated logging camps in Burma (complete descriptions will be published elsewhere and can, as well, be accessed at www.is.wayne.edu/mnissani/ ElephantCorner/do elephants know that people see.htm) resorted to numerous controls and variations. These experiments lent support to previous studies of chimpanzees and elephants. Here too elephants performed well in the three background conditions and in some experimental conditions. They performed at about chance level in other conditions, even though these conditions were conceptually similar to the others and did not require greater visual acuity.

The statistically significant performance in some conditions can perhaps be best viewed as the formation, over a lifetime, of weak associations between human faces and rewards. Hence, although the overall results do not rule out thinking and a theory of mind, to this writer they appear more compatible with the notion that elephants do not know that people see, and, more generally, that elephants lack a theory of mind.
Retrieving distant objects with a stick
The spontaneous retrieval with a stick of otherwise inaccessible objects is sometimes cited as an example of insight, although this action could also arise by pure trial and error learning. Beck (1986) placed a food tray out of reach of captive hamadryas baboons, and an L-shaped rod within reach. After 11 hours, one youngster accidentally flipped the rod over the tray and, upon retrieving the rod, brought the tray within reach. Gradually and haphazardly, he learned to use it to retrieve food.
Leaving aside, however, the question of interpreting retrieval behavior, it is of interest to review the available literature. Hobhouse (1915, p. 277) reported that a single captive elephant never learned to use a stick to get a biscuit. Time after time she “would pull the stick in to her, getting the bun if it happened to be placed exactly right, and missing it if it was possible to do so. When she missed it, she got excessively annoyed with the stick, and would try to break it by stamping on it, or throw it away into her cage.” Similar negative observations were made by Rensch and Altevogt (1954) and by Benjamin L. Hart, Lynette A. Hart and Noa Pinter-Wollman (pers. comm.). On six separate days, we have likewise failed to observe this behavior in the two Asian elephants of the Detroit Zoological Institute, even after repeatedly demonstrating to them the manipulation of food with a stick.
The one positive report known to me (Chadwick 1992) gives no details, and it is not clear whether, in this case, the behavior was directly made by the author, nor whether it could be attributed to training.
Looking back: Do elephants think?
Despite a century of intense scientific work, we still do not know if some animals, besides ourselves, think. In particular, the emerging psychological literature is more compatible with the view that elephants do not think. Elephants learn unfamiliar tasks in a mechanical, gradual fashion, with no clear evidence of comprehension. They require many trials to acquire a task, and, when the nature of the task is slightly changed (e.g., placing the lid alongside the bucket), they continue to act mechanically, suggesting that they do not understand the setup any better than Fabre’s digging wasps or Thorndike’s cats. Extensive efforts to come up with unequivocal demonstrations of insightful behavior or of theory of mind in this species are inconclusive (cf. Jayewardene 1994). On a few occasions, when seemingly insightful behavior is subject to transferability tests, the elephants fail to pass the new tests. Although uncertainties remain, two of nine elephants are perhaps capable of self-referential behavior in front of a mirror, but the meaning of such behavior remains unclear. Moreover, it is hard to believe that a minority of elephants are capable of self-recognition, while the majority is not. In short-term memory experiments, elephants never devise a rule of thumb to improve their performance.
Day et al. (2001) argue that “the evidence of high cognitive abilities in cetaceans does not stand up to close scrutiny under the standards established by laboratory researchers. This is likely to lead to a sterile debate between laboratory and field researchers unless fresh ways of taking the debate forward are found.” Needless to say, this review discloses an almost identical dichotomy in our perceptions of elephants, a dichotomy, which likewise requires fresh ways of thinking. Until such new trails are blazed, we may perhaps conclude with the following statement: while our own extensive series of controlled experiments with elephants do not rule out the widespread view that elephants think, they appear more compatible with the view that they do not.
Acknowledgements
The Detroit Zoological Institute, Myanmar Department of Forestry, the Myanma Timber Enterprise, former Ambassador U Linn Myaing, made our elephant research possible. My wife Donna shared with me her love for elephants and the joys and liverbreaks of fi eldwork. My heartfelt thanks to Dr. Wan Htun, U Tin Lay, and U Kyaw Kyaw, for guidance, guardianship, friendship, and competent assistance; may the needless clouds that have been darkening every aspect of their lives for decades disperse soon.
References
Altevogt, R. (1990) Proboscideans. In: Grzimek’s Encyclopedia of Mammals, Volume. 4. McGraw-Hill, New York.
Anderson, J.R. (1996) Chimpanzees and capuchin monkeys: Comparative Cognition. In: Reaching into Thought. Russon, A.E., Bard, K.A., Parker, S.T. (eds.) Cambridge University Press, Cambridge. pp 23-56.
Beck, B.B. (1986) Tradition and social learning in animals. In: Animal Intelligence: Insights into the Animal Mind. Hoage, R.J., Goldman, L. (eds.) Smithsonian Institution Press, Washington DC. pp 149-163.
Chadwick, D.H. (1992) The Fate of the Elephant. Sierra Club, San Francisco.
Dawkins, M. (1993) Through Our Eyes Only? W.H. Freeman, Oxford.
Day, R.L., Kendal, J.R., Kevin, N. & Laland, K.N. (2001) Validating cultural transmission in cetaceans. Behavioral and Brain Sciences 24: 330–331.
Fabre, J.H. (1915) The Hunting Wasps. Translated by Alexander Teixiera de Mattos. Dodd, New York.
Grzimek, B. (1944) Gedächtnisversuche mit Elefanten (Abstract only: Memory experiments with elephants). Zeitschrift für Tierpsychologie 6: 126-140.
Heinrich, B. (1999) Testing insight in ravens. In: The Evolution of Cognition. Heyes, C. & Huber L. (eds.) MIT, Cambridge, MA. pp 289-305.
Hobhouse, L.T. (1915) Mind in Evolution. Macmillan, London.
Jayewardene, J. (1994) The Elephant in Sri Lanka. The Wildlife Heritage Trust, Colombo, Sri Lanka.
Mackintosh, N.J., Wilson, B. & Boakes, R.A. (1985) Differences in mechanisms of intelligence among vertebrates. In: Animal Intelligence. Weiskrantz, L. (ed.) Clarendon Press, Oxford. pp 53-65.
Nissani, M. (2004) Theory of mind and insight in chimpanzees, elephants, and other Animals? In: Comparative Vertebrate Cognition. Rogers, L.J., Kaplan, G. (eds.) Kluwer Academic / Plenum Publishers, New York. pp 227-261.
Nissani, M. (2006) Do Asian elephants apply causal reasoning to tool use tasks? Journal of Experimental Psychology: Animal Behavior Processes 31: 91-96.
Nissani, M, & Hoefler-Nissani, D.M. (2004) The Inner Life of Elephants (a film) Wayne State University, Detroit. (http://youtube.com/ watch?v=YMm7j8QETi8)
Nissani, M., Hoefler-Nissani, D., Lay, U.T. & Htun, U.W. (2005) Simultaneous visual discrimination in Asian elephants. Journal of the Experimental Analysis of Behavior 83: 15-29.
Nissani, M. & Hoefler-Nissani, D. (2006) Absence of mirror self-referential behavior in two Asian elephants. J Med Biol Sci (online).
Povinelli, D.J. (1989) Failure to find self-recognition in Asian elephants (Elephas maximus) in contrast to their use of mirror cues to discover hidden food. Journal of Comparative Psychology 103: 122-131.
Povinelli, D.J. & Eddy, T.J. (1996) What young chimpanzees know about seeing. Monographs of the Society for Research in Child Development 61(3): 1-189.
Rensch, B. (1957) The intelligence of elephants. Scientific American 196: 44-49.
Rensch, B. & Altevogt, R. (1954) Zähmung und Dressurleistungen indischer Arbeitselefanten (Translated title: Taming and training performances of Indian work elephants) Zeitschrift für Tierpsychologie 11: 497-510.
Simonet, P. (2000) Self-recognition in Asian elephants, preliminary findings. Elephant 2: 103.
Simonet, P., Myers, M, Lamoureux L, Quashnick J, Jacobs J & Willison K (2000) Social and cognitive factors in Asian elephant mirror behavior & self-recognition. Unpublished manuscript.
Stevens, V.J. (1978) Basic operant research in the zoo. In: Behavior of Captive Wild Animals. Markowitz, H. & Stevens, V.J. (eds.) Nelson-Hall, Chicago.
Swartz, K.B. & Evans, S. (1991) Not all chimpanzees (Pan troglodytes) show self-recognition. Primates 32: 483-496.
Thorpe, W.H. (1956) Learning and Instinct in Animals. Harvard University Press, Cambridge, MA.
Thorndike, E.L. (1911) Animal Intelligence. Macmillan, New York.
Williams, J.H. (1950) Elephant Bill. Doubleday, Garden City.
Author’s e-mail: moti.nissani@wayne.edu